Le monochromatisme à cônes bleus est une maladie génétique rare de la rétine causée par des mutations génétiques sur les gènes OPN1LW, OPN1MW et LCR. Ces gènes sont localisés sur le chromosome X, et codent pour des protéines appelées photopigments, nécessaires aux cônes rouges et verts pour capturer la lumière.
Nous verrons sur cette page quels sont les gènes responsables de la vision de la couleur, quelles sont les mutations génétiques qui mènent à la maladie et l’histoire de ces découvertes scientifiques de génétique moléculaire.
L’association BCM Families Foundation soutient la recherche de génétique moléculaire sur le monochromatisme à cônes bleus et la création d’un Registre des Patients souffrant de monochromatisme à cônes bleus, en vue d’approfondir les connaissances sur les mutations génétiques qui sont à l’origine de la maladie.
Les gènes responsables de la vision de la couleur
Le monochromatisme à cônes bleus est causé par des mutations génétiques sur certains gènes qui se trouvent sur le chromosome X.
Le chromosome X est composé de deux bras: le bras supérieur (appelé p), le bras inférieur (q).
Les gènes impliqués dans le monochromatisme à cônes bleus se trouvent dans la position Xq28, à la fine du bras q.
La figure ci-après montre le chromosome X.
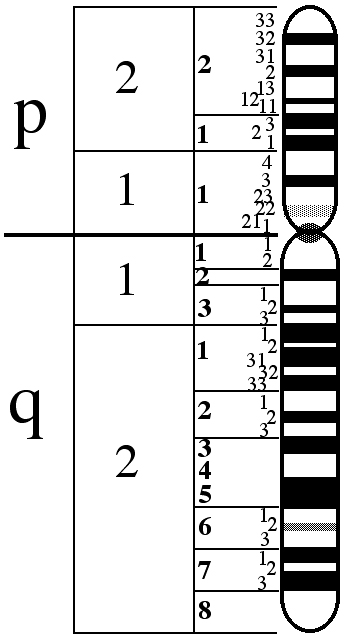
Dans la position Xq28 se trouvent, dans l’ordre, les gènes identifiés par les noms LCR, OPN1LW et OPN1MW.
LCR (du nom anglais Locus Control Région) est la région de contrôle du locus et agit comme promoteur de l’expression des deux gènes suivants. En l’absence de ce gène, aucun des deux gènes suivants n’est exprimé dans la rétine humaine.
En outre, la LCR assure qu’un seul des deux gènes opsine (rouge ou vert) soit exprimé de manière exclusive dans chaque cône.
OPN1LW et OPN1MW sont les gènes qui contiennent le code génétique pour les protéines opsines, c’est-à-dire les protéines qui constituent les photopigments pour la capture de la lumière, respectivement de couleur rouge (Long Wave = ondes longues) et de couleur verte (Medium Wave = ondes moyennes).
De nombreuses personnes ont plusieurs répétitions du gène relatif au photopigment vert, OPN1MW. Toutefois, seuls les deux premiers gènes, tout de suite après la LCR, sont exprimés dans la rétine, tandis que les répétitions suivantes ne sont pas exprimées. Par exemple, un homme caucasien aura dans 25% des cas un seul gène OPN1MW, dans 50% des cas il en aura 2 et dans les 25% restants, il en aura 3 ou plus.
Dans la figure ci-dessous, nous voyons une représentation de ces gènes lorsqu’il y a absence de mutations génétiques pathologiques.
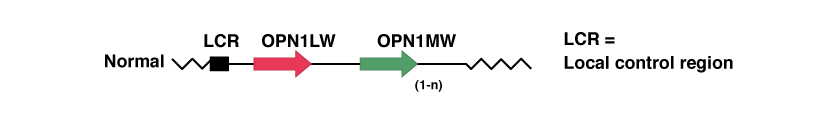
Pour approfondir la connaissance de ces gènes, il est possible de consulter le site du «National Center for Biotechnology Information», NCBI:
- OPN1LW, photopigment de cône rouge;
- OPN1MW, photopigment de cône vert;
- LCR, région de contrôle du locus.
Le gène responsable de la formation du photopigment bleu se trouve dans une position très éloignée, dans le chromosome 7, et le gène responsable de la formation de la rhodopsine (le photopigment des bâtonnets) se trouve sur le chromosome 3:
- OPN1SW photopigment de cône bleu;
- RHODOPSINE photopigment des bâtonnets.
Dans la figure suivante, nous voyons les protéines opsines des cônes bleus «S» (short = court), verts «M» (medium = moyen) et rouges «L» (long).
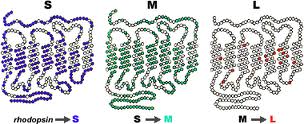
(La figure est tirée du site handprint.com)
Elles ont la forme d’une chaîne qui passe 7 fois à travers un disque du segment externe d’un cône. Les trois protéines sont très semblables entre elles et, en particulier, la M et la L diffèrent seulement par certains éléments. Les deux photopigments, le rouge et le vert, sont en effet égaux à 96%, tandis qu’ils ont une ressemblance de 46% seulement avec le photopigment bleu.
Les gènes OPN1LW et OPN1MW, comme tous les gènes, sont formés d’exons et d’introns. Plus précisément, ces gènes possèdent 6 exons, numérotés de 1 à 6.
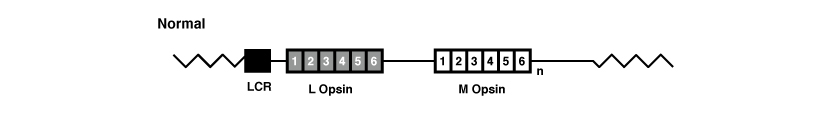
(La figure est tirée de l’ouvrage de Jessica C. Gardner, Michel Michaelides, Graham E. Holder, Naheed Kanuga, Tom R. Webb, John D. Mollon, Anthony T. Moore, Alison J. Hardcastle ‘Blue cone monochromacy: Causative mutations and associated phenotypes’ Molecular Vision 2009; 15:876-884).
Comme toutes les protéines, les opsines sont elles aussi des structures tridimensionnelles qui doivent effectuer un «repliement» (en anglais: folding) pour acquérir leur forme tridimensionnelle finale. Certains aminoacides présents à l’intérieur de la protéine sont responsables de ce repliement.
Les mutations génétiques
Il existe de nombreuses mutations génétiques susceptibles de frapper ce groupe de gènes: LCR, OPN1LW et OPN1MW.
Certaines mutations mènent des à conditions communément désignées sous le nom de «daltonisme» et dont le seul effet est l’impossibilité de distinguer certaines couleurs.
Les mutations menant au monochromatisme à cônes bleus identifiées jusqu’ici sont les suivantes :
Grandes délétions
Délétion de la LCR, ou délétion de la LCR et d’une partie ou de la totalité des exons du gène OPN1LW.
Cette mutation correspond à l’absence d’une grande partie du matériel génétique. Le code génétique LCR n’étant pas présent, les deux protéines ne seront pas exprimées et les cônes seront donc dépourvus des photopigments rouge et vert.
Délétion intragénique. Il s’agit d’une délétion d’exons à l’intérieur des gènes OPN1LW et OPN1MW ou d’une délétion du matériel génétique du premier gène et d’une partie du second.
Cette mutation correspond elle aussi à l’absence d’une grande partie du matériel génétique.
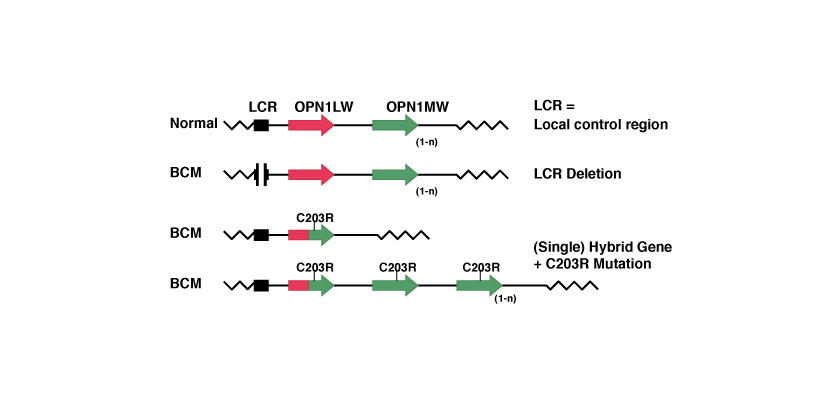
Mécanisme à 2 étapes avec recombinaison homologue et inactivation.
Dans ce cas, compte tenu de la ressemblance entre les deux gènes OPN1LW et OPN1MW, lors d’un processus de «recombinaison homologue» l’un des deux gènes se perd et il se crée un nouveau gène, qui est hybride. Ensuite, une mutation ponctuelle rend inactif le gène qui est resté.
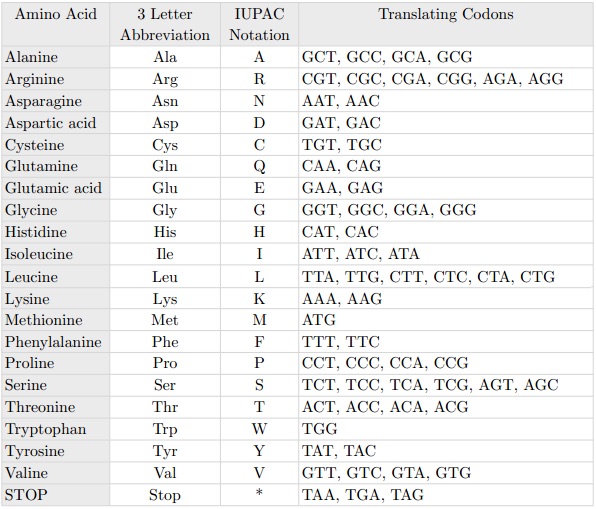
La mutation ponctuelle la plus connue est celle qui porte le nom de C203R. Le nom des mutations ponctuelles indique la position à laquelle est survenue la mutation (ici, la position 203) et l’aminoacide qui a été remplacé (ici, la cystéine C, remplacée par l’arginine R). Au niveau des codons, cette substitution est ponctuelle parce qu’elle équivaut à remplacer la thymine par la cytosine dans la position 648, comme nous pouvons le voir sur le tableau suivant:
La mutation C203R empêche la protéine opsine, une fois codée, de se replier dans sa forme tridimensionnelle.
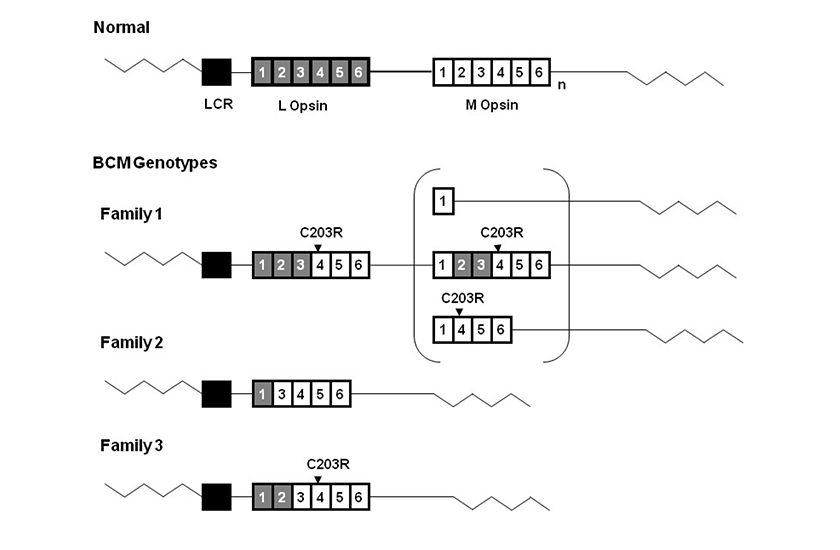
(La figure est tirée de l’ouvrage de Jessica C. Gardner, Michel Michaelides, Graham E. Holder, Naheed Kanuga, Tom R. Webb, John D. Mollon, Anthony T. Moore, Alison J. Hardcastle ‘Blue cone monochromacy: Causative mutations and associated phenotypes’ Molecular Vision 2009 ; 15 :876-884).
Génotypes du monochromatisme à cônes bleus. Diagramme représentant les génotypes du monochromatisme à cônes bleus de 3 familles britanniques. La matrice opsine L-M de type sauvage est visible au sommet de la figure. Les cases grises représentent les exons d’opsine M. L’indice « n » représente un ou plusieurs gènes d’opsine M. La case noire représente la région de contrôle du locus LCR. La LCR était présente sans mutations dans les trois familles. Les mutations ponctuelles C203R détectées dans les familles 1 et 3 sont visibles au-dessus des exons correspondants. La famille 1 a un gène hybride inactif suivi d’un second gène dans la matrice. On peut voir entre parenthèses trois structures possibles de ce second gène inactif. La famille 2 a un seul gène hybride défectueux, dépourvu de l’exon 2. La famille 3 a un seul gène hybride inactif.
D’autres mutations ponctuelles sont les mutations P307L et R247X. La dernière remplace l’arginine par le codon STOP, à la position 247, ce qui empêche la formation de la protéine (mutation non-sens).
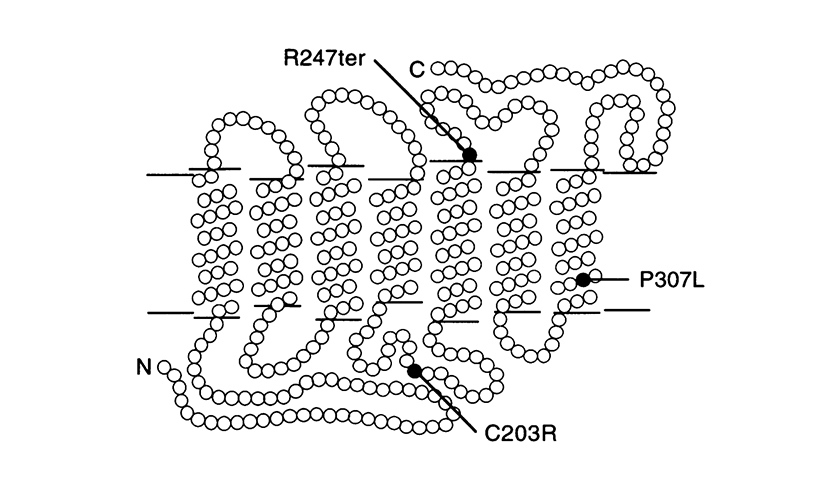
(La figure est tirée de l’ouvrage de J. Nathans et al. Am. J. Hum. Genet. 53 : 987-1000, 1993).
Modèle de pigment rouge et de pigment hybride 5’ rouge – 2’ vert dans la membrane du photorécepteur, montrant les emplacements des mutations ponctuelles identifiées chez les patients atteints de monochromatisme à cônes bleus. Chaque cercle représente un aminoacide. « N » = terminaison amine et « C » = terminaison carboxyle. La terminaison amine fait face à l’espace extracellulaire.
Autres mutations
D’autres mutations sur les gènes OPN1LW et OPN1MW menant au monochromatisme à cônes bleus sont constituées par un ensemble de mutations ponctuelles, appelées par exemple LIAVA. La personne aura le monochromatisme à cônes bleus si, comme dans le cas précédent, il se produit, par effet d’une recombinaison homologue, un unique gène mixte entre l’OPN1LW et l’OPN1MW et que l’exon 3 contient les aminoacides suivants dans les positions indiquées: Leucine 153, Isoleucine 171, Alanine 174, Valine 178 et Alanine 180. Ce génotype porte le nom abrégé de LIAVA.
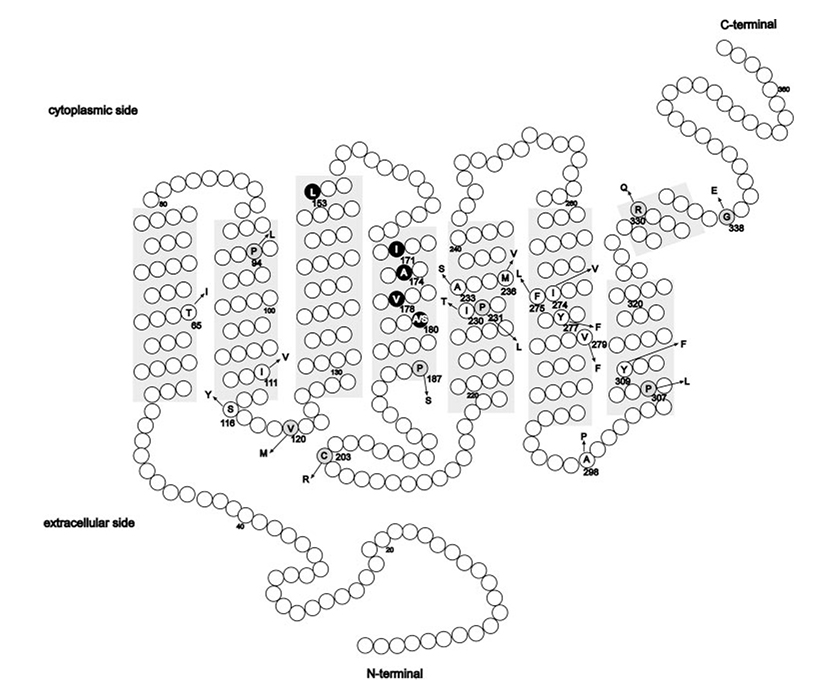
(La figure est tirée de l’ouvrage de Mizrahi-Meissonnier L., Merin S., Banin E., Sharon D., 2010).
Emplacement des altérations des aminoacides reportées jusqu’ici dans les gènes d’opsine L et M. Zones ombrées: domaines transmembranaires. Cercles : différences entre aminoacides et polymorphisme connu avec l’aminoacide le plus commun (indiqué par un code à une lettre). Flèche: modification de l’aminoacide. Le nombre de codons est décrit pour chaque modification. Les changements faux-sens, associés à une maladie des cônes liée à l’opsine, susceptibles de causer des dysfonctionnements protéiniques sont indiqués sur fond gris. L’haplotype LIAVA est surligné en noir.
Nathans et al. 1993 | ||
Ayyagari et al. 2000 | ||
A.Reitner, L.T. Sharpe, E. Zrenner 1991 | ||
Mizrahi-Meissonnier L. et al. 2010 Neitz M. et al. 2004 Crognale MA. et al. 2004 |
Autres maladies comportant des mutations génétiques sur les gènes OPN1LW et OPN1MW
Une autre maladie de la rétine associée à la position Xq28 est la maladie oculaire de Bornholm (BED – Bornholm Eye Disease), dont les symptômes sont similaires à ceux du monochromatisme à cônes bleus. Il s’agit d’une maladie très rare et stationnaire. Pour approfondir, on peut consulter OMIM et le site de l’Université de l’Arizona.
Enfin, rappelons qu’il existe également une mutation particulière des deux gènes OPN1LW et OPN1MW qui cause une maladie différente du monochromatisme à cônes bleus. Il s’agit de la mutation de type «mauvais repliement» (en anglais: misfolding), W177R, qui, si elle est présente sur les deux gènes opsines, provoque la dystrophie des cônes avec évidence de dégénération et mort des cellules des cônes.
Histoire de la découverte des gènes du monochromatisme à cônes bleus
De nombreux chercheurs ont contribué aux découvertes concernant les gènes impliqués dans le monochromatisme à cônes bleus.
Rappelons les découvertes fondamentales de Jeremy Nathans sur les gènes responsables de la vision de la couleur:
Nathans, J., Thomas, D., Hogness, D. S. Molecular genetics of human color vision: the genes encoding blue, green, and red pigments. Science 232: 193-202, 1986. PMID:2937147.
Nathans, J., Piantanida, T. P., Eddy, R. L., Shows, T. B., Hogness, D. S. Molecular genetics of inherited variation in human color vision. Science 232: 203-210, 1986. PMID: 3485310 .
Nathans, J. Molecular biology of visual pigments. Annu. Rev. Neurosci. 10: 163-194, 1987. PMID: 3551758 .
Nathans, J. The evolution and physiology of human color vision: insights from molecular genetic studies of visual pigments. Neuron. 24: 299-312, 1999. PMID: 10571225 .
Deeb, S. S. The molecular basis of variation in human color vision. Clin. Genet. 67: 369-377, 2005. PMID: 15811001 .
En particulier, les travaux qui ont permis de comprendre les principales causes du monochromatisme à cônes bleus et notamment le processus à 2 étapes avec mutation ponctuelle C2013R:
Nathans, J., Davenport, C. M., Maumenee, I. H., Lewis, R. A., Hejtmancik, J. F., Litt, M., Lovrien, E., Weleber, R., Bachynski, B., Zwas, F., Klingaman, R., Fishman, G. Molecular genetics of human blue cone monochromacy. Science 245: 831-838, 1989. PMID: 2788922.
Nathans, J., Maumenee, I. H., Zrenner, E., Sadowski, B., Sharpe, L. T., Lewis, R. A., Hansen, E., Rosenberg, T., Schwartz, M., Heckenlively, J. R., Traboulsi, E., Klingaman, R., Bech-Hansen, N. T., LaRoche, G. R., Pagon, R. A., Murphey, W. H., Weleber, R. G. Genetic heterogeneity among blue-cone monochromats. Am. J. Hum. Genet. 53: 987-1000, 1993. PMID: 8213841.
Reyniers, E., Van Thienen, M.-N., Meire, F., De Boulle, K., Devries, K., Kestelijn, P., Willems, P. J. Gene conversion between red and defective green opsin gene in blue cone monochromacy. Genomics 29: 323-328, 1995. PMID: 8666378.
Un travail important pour les mutations de type «Délétion» de la LCR ou de la LCR et d’une partie du gène OPN1LW est:
Ayyagari, R., Kakuk, L. E., Bingham, E. L., Szczesny, J. J., Kemp, J., Toda, Y., Felius, J., Sieving, P. A. ‘Spectrum of color gene deletions and phenotype in patients with blue cone monochromacy’. Hum. Genet. 107: 75-82, 2000. Hum Genet. 2000 Jul;107(1):75-82. PMID: 10982039.
Pour la délétion intragénique, rappelons les ouvrages suivants, qui identifièrent un cas de monochromatisme à cônes bleus avec présence du seul gène OPN1LW du photopigment rouge, mais avec absence de l’exon 4 de ce gène:
Ladekjaer-Mikkelsen, A.-S., Rosenberg, T., Jorgensen, A. L. ‘A new mechanism in blue cone monochromatism’. Hum. Genet. 98: 403-408, 1996. PMID: 8792812.
Reitner, A., Sharpe, L. T., Zrenner, E. Is colour vision possible with only rods and blue-sensitive cones? Nature 352: 798-800, 1991. PMID: 1881435 .
La LCR, et son rôle dans l’expression des gènes de l’opsine, est le résultat des travaux suivants:
Lewis, R. A., Holcomb, J. D., Bromley, W. C., Wilson, M. C., Roderick, T. H., Hejtmancik, J. F. Mapping X-linked ophthalmic diseases: III. Provisional assignment of the locus for blue cone monochromacy to Xq28. Arch. Ophthal. 105: 1055-1059, 1987. PMID: 2888453 .
Lewis, R. A., Nathans, J., Holcomb, J. D., Bromley, W. C., Roderick, T. H., Wilson, M. C., Hejtmancik, J. F. ‘Blue cone monochromacy: assignment of the locus to Xq28 and evidence for its molecular rearrangement’. Am. J. Hum. Genet. 41: A102 only, 1987.
Wang, Y., Macke, J. P., Merbs, S. L., Zack, D. J., Klaunberg, B., Bennett, J., Gearhart, J., Nathans, J. ‘A locus control region adjacent to the human red and green visual pigment genes’. Neuron 9: 429-440, 1992. PMID: 1524826.
En particulier, la fonction permettant l’expression exclusive d’une opsine (rouge ou verte) dans chaque cône a été découverte dans le dernier ouvrage.
Pour l’étude de la mutation C203R, rappelons:
Kazmi MA, Sakmar TP, Ostrer H. ‘Mutation of a conserved cysteine in the X-linked cone opsins causes color vision deficiencies by disrupting protein folding and stablilty’. Investigative Ophthalmology and Visual Science. 1997; 38(6):1074–1081.
qui comprirent les effets négatifs de cette mutation sur le repliement des opsines et:
Winderickx J, Sanocki E, Lindsey DT, Teller DY, Motulsky AG, Deeb SS. Defective colour vision associated with a missense mutation in the human green visual pigment gene. Nature Genetics. 1992;1:251–256. PMID: 1302020 .
qui étudièrent cette mutation et sa fréquence d’environ 2% chez les personnes d’origine caucasienne.
Sur les mutations rares du type LIAVA, l’on peut consulter:
Carroll J1, Neitz M, Hofer H, Neitz J, Williams DR., ‘Functional photoreceptor loss revealed with adaptive optics: an alternate cause of color blindness.’ Proc Natl Acad Sci U S A. 2004 Jun. PMID: 15148406.
Mizrahi-Meissonnier L., Merin S., Banin E., Sharon D., ‘Variable retinal phenotypes caused by mutations in the X-linked photopigment gene array’. Invest. Ophthalmol. Vis. Sci. 2010 Aug;51(8):3884-92. PMID: 20220053.
Neitz M, Carroll J, Renner A, et al. ‘Variety of genotypes in males diagnosed as dichromatic on a conventional clinical anomaloscope’. Vis Neurosci. 2004;21:205–216. PMID: 15518190.
Crognale MA, Fry M, Highsmith J, et al. ‘Characterization of a novel form of X-linked incomplete achromatopsia’. Vis Neurosci. 2004; 21:197–203. PMID: 15518189.
Certains ouvrages historiques pendant les premiers stades de la recherche sur le monochromatisme à cônes bleus ont été:
Huddart, J. ‘An account of persons who could not distinguish colours’. Phil. Trans. Roy. Soc. 67: 260 only, 1777.
Sloan, L. L. ‘Congenital achromatopsia: a report of 19 cases’. J. Ophthal. Soc. Am. 44: 117-128, 1954. PMID: 13131176.
Alpern, M., Falls, H. F., Lee, G. B. ‘The enigma of typical total monochromacy’. Am. J. Ophthal. 50: 996-1012, 1960. PMID: 13682677.
Blackwell, H. R., Blackwell, O. M. Rod and cone receptor mechanisms in typical and atypical congenital achromatopsia. Vision Res. 1: 62-107, 1961. https://doi.org/10.1016/0042-6989(61)90022-0.
Fleischman, J. A., O’Donnell, F. E. Jr. ‘Congenital X-linked incomplete achromatopsia. Evidence for slow progression, carrier fundus findings, and possible genetic linkage with glucose-6-phosphate dehydrogenase locus’. Arch Ophthalmol 1981;99:468-472. PMID: 6971088.
Lewis, R. A., Holcomb, J. D., Bromley, W. C., Wilson, M. C., Roderick, T. H., Hejtmancik, J. F. ‘Mapping X-linked ophthalmic diseases: III. Provisional assignment of the locus for blue cone monochromacy to Xq28′. Arch. Ophthal. 105: 1055-1059, 1987. PMID: 2888453.
Pour l’étude de la dystrophie des cônes, une maladie dégénérative due à une mutation ponctuelle sur les deux gènes OPN1LW et OPN1MW:
Gardner JC, Webb TR, Kanuga N, Robson AG, Holder GE, Stockman A, Ripamonti C, Ebenezer ND, Ogun O, Devery S, Wright GA, Maher ER, Cheetham ME, Moore AT, Michaelides M and Hardcastle AJ,’X-Linked Cone Dystrophy Caused by Mutation of the Red and Green Cone Opsins’.The American Journal of Human Genetics 87, 26–39, July 9, 2010. PMID: 20579627.
Reportons également certains articles de revues qui illustrent le sujet:
Neitz J., Neitz M. ‘The genetics of normal and defective color vision’. 2011 Review. Vision Research. PMID: 21167193.
Deeb, S.S. ‘Molecular Genetics of colour vision deficiencies’. Clinical and Experimental Optometry 87.4 – 5 July 2004. PMID:15312026.
